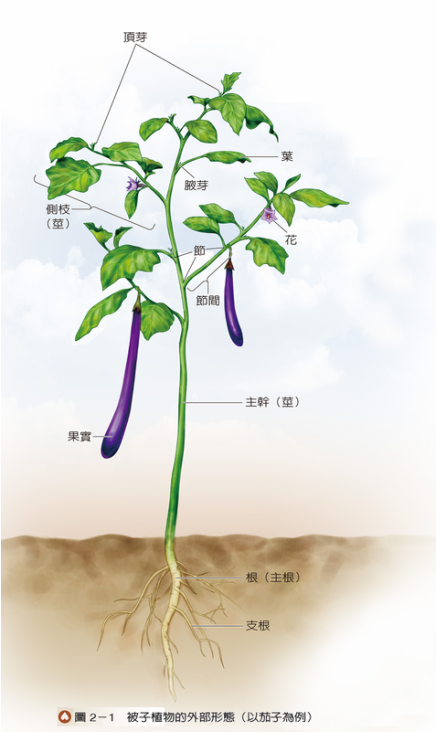
在各種植物當中,維管束植物具有特化的維管束,是植物體輸送物質的主要管道,也具有支持的功能,故個體相對較為高大。根、莖、葉是植物體吸收、運輸或合成營養的主要構造,屬於維管束植物的營養器官。以下將以被子植物(圖2-1)為例,分別介紹根、莖、葉的形態、構造與功能。

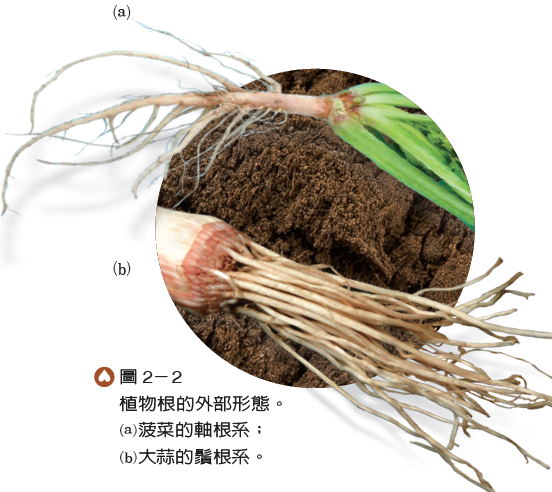
根通常是植物體的地下部分,外形多細長、有些可長出支根,但不具節,且沒有葉著生其上。根可固定和支持植物體,並從土壤中吸收水和無機鹽。此外,根中的細胞還具有儲存水和養分的功能。

植物體所有的根總稱為根系,通常分為軸根系和鬚根系兩種類型(圖2-2)。根系中可明顯分出主根、支根的粗細差異者,稱為軸根系,如一般雙子葉植物的根系。雙子葉植物的種子萌發時,先長出初生根,然後初生根會發育成主根,再由主根分生出許多較細的支根,而構成軸根系。鬚根系則是由許多粗細相似的根所構成,一般單子葉植物的根系屬之。單子葉植物的初生根存活一段時間後,即會停止生長或枯萎消失,並在幼莖的基部長出許多不定根,而形成粗細相似的根,構成鬚根系。

不定根
典型的根是由初生根發育而來的。然而,有些植物尚可直接從莖或葉產生新根,由於這些根不是源自根部組織,故稱為不定根。除了組織起源不同之外,不定根與一般的根具有相似的構造與功能,可吸收、運輸水分及無機鹽與協助支持植物體;但不定根可能還有其他特殊的功能,例如:榕樹(下圖)莖上有鬚狀的根懸垂在空中,稱為氣生根,可直接吸收空氣中的氧及水分。這些氣生根可持續生長至接觸地面,然後伸入土中,協助吸收水分,並加粗如同許多小「樹幹」般,以支持植物體。此時,這些粗壯的不定根改稱為支持根。
典型的根是由初生根發育而來的。然而,有些植物尚可直接從莖或葉產生新根,由於這些根不是源自根部組織,故稱為不定根。除了組織起源不同之外,不定根與一般的根具有相似的構造與功能,可吸收、運輸水分及無機鹽與協助支持植物體;但不定根可能還有其他特殊的功能,例如:榕樹(下圖)莖上有鬚狀的根懸垂在空中,稱為氣生根,可直接吸收空氣中的氧及水分。這些氣生根可持續生長至接觸地面,然後伸入土中,協助吸收水分,並加粗如同許多小「樹幹」般,以支持植物體。此時,這些粗壯的不定根改稱為支持根。


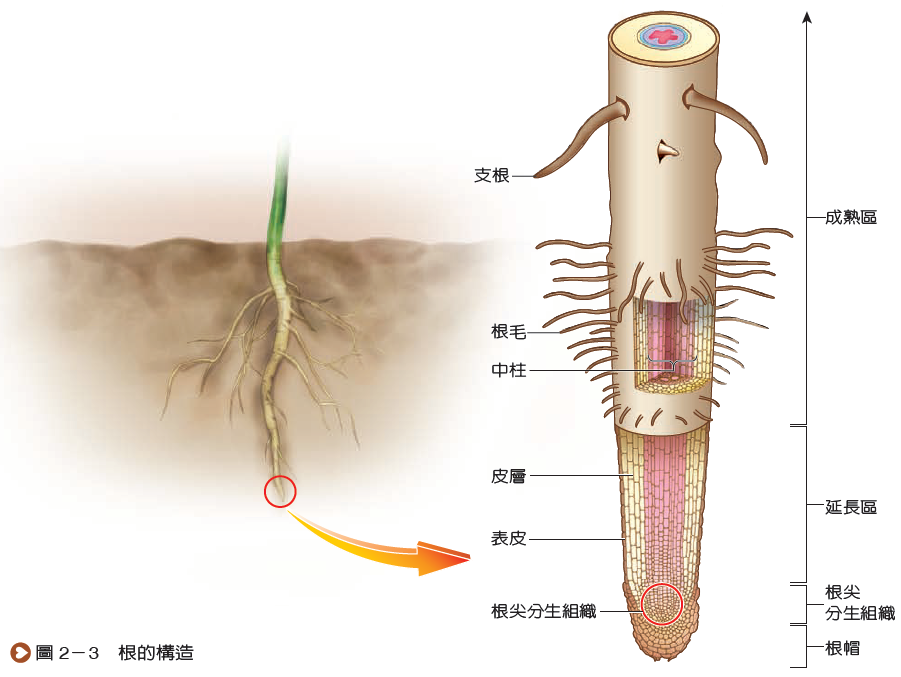
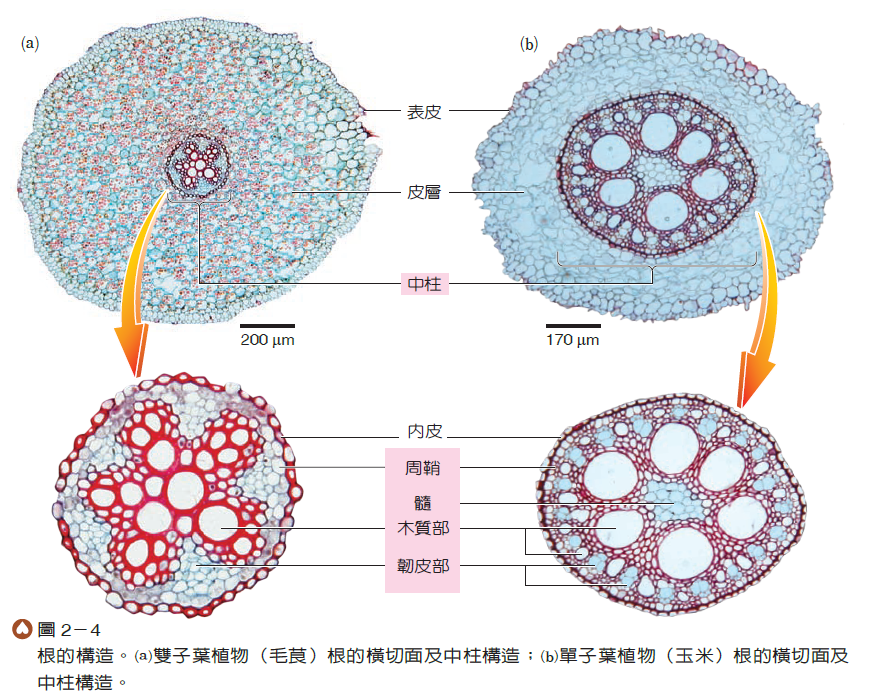
大多數植物的根由先端往上依次可區分為根帽、根尖分生組織、延長區和成熟區四部分(圖2-3)。根帽位於根的最先端,又稱根冠,覆蓋根尖分生組織,可保護根尖在穿透土壤時免於受損。根尖分生組織的細胞分裂最頻繁,可使根的主軸持續延伸,利於擴大根系的範圍。分生組織產生的新細胞,一部分補充脫落的根帽細胞,大部分繼續特化為組成根的細胞,這些細胞通常會長大至成熟細胞的大小,並逐漸表現出該細胞的特殊功能,故此部位稱為延長區。成熟區的細胞大多不再繼續長大,且與其他特化的細胞構成不同的構造,由外而內分為表皮、皮層和中柱三部分(圖2-4),彼此分工合作。
點圖有切片資料
根的表皮位於最外層,具有保護及吸收的功能。在剛成熟的根中,有些表皮細胞會局部向外凸出而形成根毛,可增加吸收的表面積,是植物體吸收水和無機鹽的主要區域。當遠離根尖或在較粗的根中,其表皮則較少凸出,雖仍可吸收水分,但逐漸以保護功能為主。
根的皮層細胞排列較為疏鬆,具有儲存的功能。皮層最內側常有一層排列緊密的細胞,稱為內皮,具管制水和無機鹽進入中柱的功能。
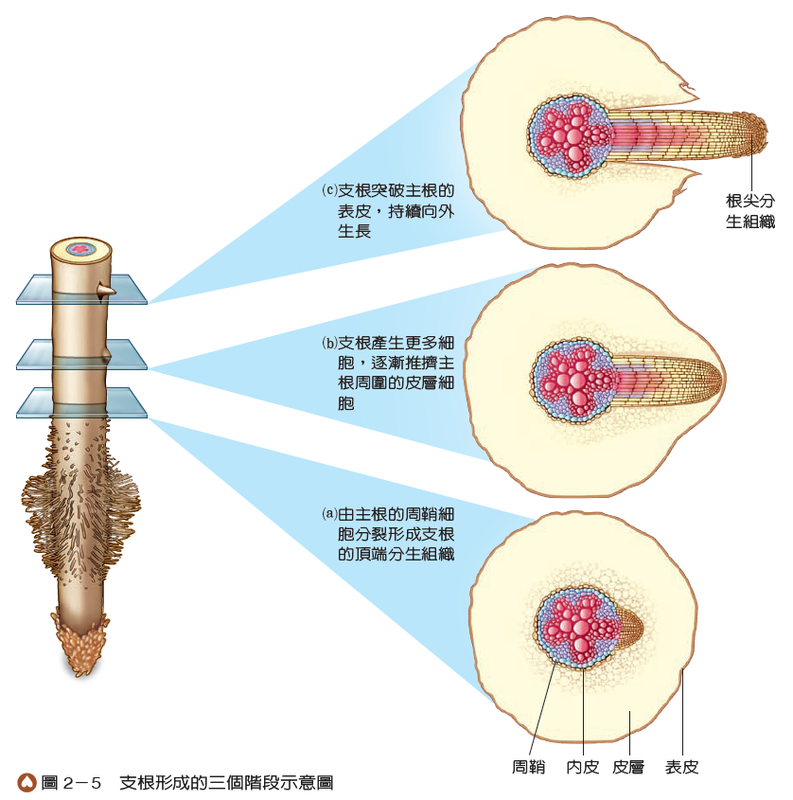
內皮以內的部分統稱為中柱,包括周鞘和維管束。周鞘位於中柱的外層,其細胞仍具有分裂的能力,支根即由此處產生(圖2-5)。周鞘內側有木質部和韌皮部,兩者常呈放射狀交錯排列,分別具有輸送水分和養分的功能。雙子葉植物與單子葉植物根的組成大致相似,但大部分單子葉植物根在中柱的中央還具有由薄壁細胞組成的髓(圖2-4(b)),而雙子葉植物較少有此構造。
根的皮層細胞排列較為疏鬆,具有儲存的功能。皮層最內側常有一層排列緊密的細胞,稱為內皮,具管制水和無機鹽進入中柱的功能。
內皮以內的部分統稱為中柱,包括周鞘和維管束。周鞘位於中柱的外層,其細胞仍具有分裂的能力,支根即由此處產生(圖2-5)。周鞘內側有木質部和韌皮部,兩者常呈放射狀交錯排列,分別具有輸送水分和養分的功能。雙子葉植物與單子葉植物根的組成大致相似,但大部分單子葉植物根在中柱的中央還具有由薄壁細胞組成的髓(圖2-4(b)),而雙子葉植物較少有此構造。

許多植物演化出不同形態與功能的變態根,以適應不同的生活環境。例如:玉米從接近地面的節長出支持根,可加強固著能力,避免被風吹倒;生存在河口沼澤的紅樹林植物海茄苳,無法從缺氧的土壤中獲得足夠的氧,因而發展出伸出地面的呼吸根;有些附著於樹幹或石壁上生長的蘭花,具有氣生根,可協助植物體附著,並吸收空氣中的水分及儲存;此外,日常食用的胡蘿蔔,其軸根儲存大量養分,屬於儲藏根(圖2-6)。


莖通常是植物體的地上部分,是支持與運輸的主要器官。自根吸收的水分與無機鹽運送至莖,再由莖運送至葉,提供光合作用之需。而葉進行光合作用產生的有機養分,亦藉由莖運送至其他部位,提供生長所需或加以儲藏。此外,有些植物的莖也具有行光合作用及儲存水分、有機養分的功能。

莖通常呈直立、具分枝的長柱形,頂端的生長區稱為頂芽,內有莖頂分生組織。莖上有節,在此處長出葉及芽;節上的芽通常位於葉腋處,故稱為腋芽(請見P.46圖2-1),可發育成新的枝條或花。
莖可依質地不同分為草本和木本兩大類。草本植物的莖大多草質多汁、柔軟易折,且多為綠色,可行光合作用,如空心菜、水稻等。木本植物的莖木質化,故支持力較佳,個體通常可持續增高和加粗。
莖的構造大致包括表皮、皮層和維管束,但雙子葉與單子葉植物莖的維管束排列方式不同,其橫向加粗的能力也有明顯的差異。
莖可依質地不同分為草本和木本兩大類。草本植物的莖大多草質多汁、柔軟易折,且多為綠色,可行光合作用,如空心菜、水稻等。木本植物的莖木質化,故支持力較佳,個體通常可持續增高和加粗。
莖的構造大致包括表皮、皮層和維管束,但雙子葉與單子葉植物莖的維管束排列方式不同,其橫向加粗的能力也有明顯的差異。

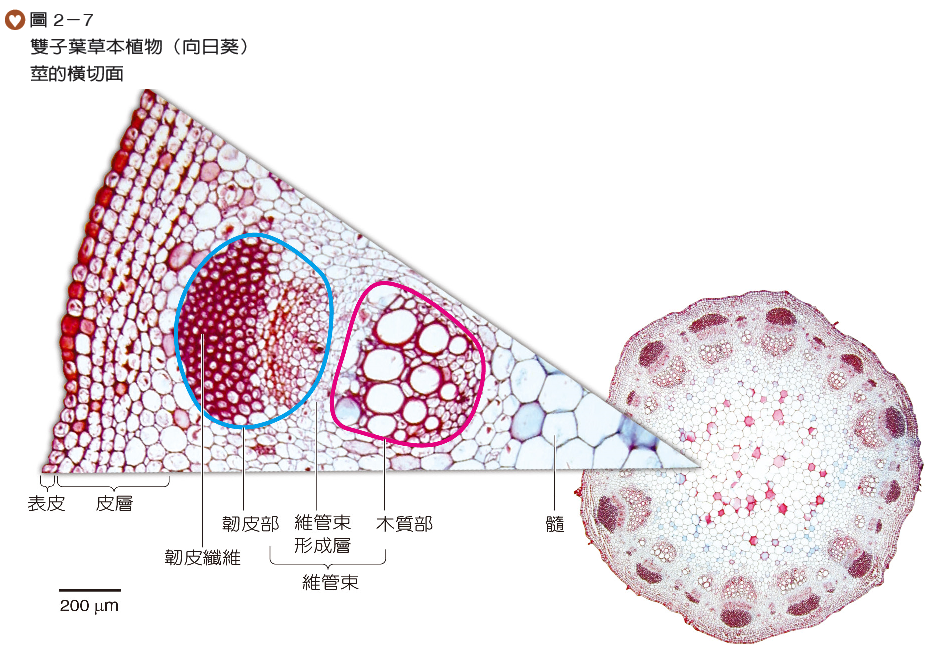
雙子葉草本植物莖的構造由外而內可區分為表皮、皮層、維管束及髓(圖2-7)。表皮由排列緊密的細胞所構成,有氣孔分布;皮層和髓主要由排列疏鬆的細胞構成,鄰近表皮的皮層細胞含有較多葉綠體,是綠色莖行光合作用的位置;維管束呈環狀排列,由韌皮部、維管束形成層與木質部組成,韌皮部位於外側,木質部位於內側。維管束形成層因受植物本身生長或壽命的限制,通常無法產生大量維管束組織,故莖直徑的增加不明顯。

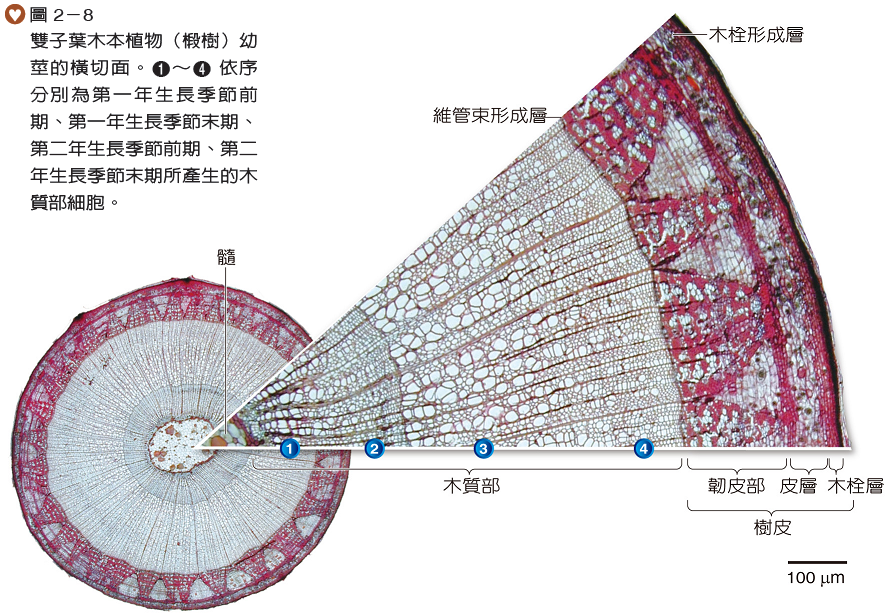
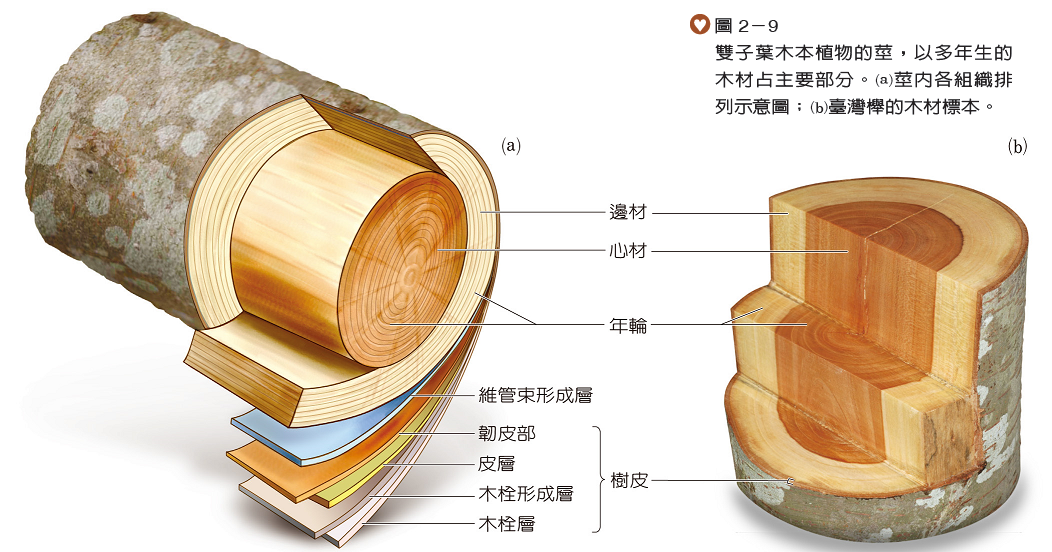
雙子葉木本植物的幼莖與草本植物的莖構造相似,由於幼莖的維管束形成層持續形成新的細胞,往外新增韌皮部,往內新增木質部,堅硬的木質部組織累積成木材,而使莖長成粗壯的木本莖(圖2-8)。
點圖至木材光顯樣本製造技術
生長於溫帶地區的樹種,其木質部細胞會隨著生長季節變換而有明顯的變化。在生長季節前期(通常在春季初),木質部的細胞長得較大且壁較薄(圖2-8、),所形成的木材顏色較淺,稱為早材;然而在生長季節末期(通常在夏季末),木質部的細胞較小且壁較厚(圖2-8、),所形成的木材顏色較深,稱為晚材,因此在樹幹木材的橫切面上,可見到顏色深淺交替形成的同心環紋,稱為年輪(圖2-9)。通常可由年輪的數量推測樹木的年齡,亦可從年輪的寬窄來推測當年的氣候狀況。
許多木本植物的莖在加粗過程中,中央的髓逐漸死亡,而外側的表皮也會被撐破,此時皮層的外層細胞會分化成木栓形成層,並向外增生形成木栓層(圖2-9(a)),以取代表皮,成為莖最外層的保護構造。木栓層不透水也不透氣,但可藉著表面常有的破裂缺口,即皮孔,來進行氣體交換(圖2-10)。此外,樹皮是木材外層組織的統稱,主要包括表皮、木栓層、木栓形成層、皮層和韌皮部等構造(圖2-9),是樹幹的保護層。
許多木本植物的莖在加粗過程中,中央的髓逐漸死亡,而外側的表皮也會被撐破,此時皮層的外層細胞會分化成木栓形成層,並向外增生形成木栓層(圖2-9(a)),以取代表皮,成為莖最外層的保護構造。木栓層不透水也不透氣,但可藉著表面常有的破裂缺口,即皮孔,來進行氣體交換(圖2-10)。此外,樹皮是木材外層組織的統稱,主要包括表皮、木栓層、木栓形成層、皮層和韌皮部等構造(圖2-9),是樹幹的保護層。


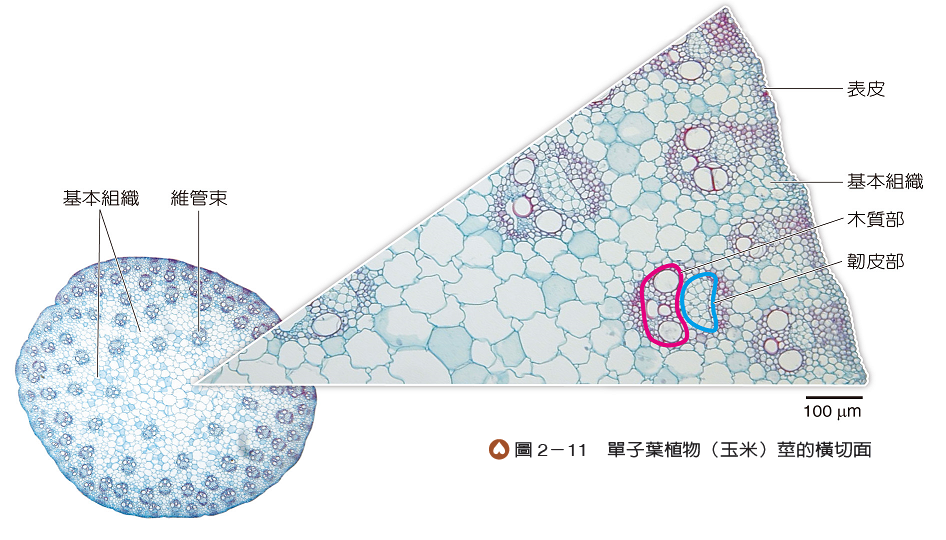
單子葉植物的莖通常沒有明顯的橫向加粗,幼莖與老莖的粗細不會差太多。構造包括表皮、基本組織及維管束(圖2-11)。維管束散布在基本組織中,且在靠近表皮處分布較密集,維管束的木質部位於內側、韌皮部位於外側,兩者之間沒有維管束形成層。

有些植物除了有典型莖的構造與功能外,還有演變成其他形式且具特殊功能的變態莖。例如:馬鈴薯的塊莖位於土壤中,可儲存養分;蛇莓的走莖橫於地表,可延伸植物體的範圍,並可在節上長出新個體;薑的根莖橫走於土壤中,可延伸植物體的範圍,並可再長出新芽,內部還儲藏大量養分;魯花樹莖的節上有「刺」,是由莖的側枝特化而成,具防禦功能(圖2-12)。

為何發芽的馬鈴薯不能吃?詳情請點圖

葉是維管束植物進行光合作用的主要部位,著生在莖的節上,壽命通常有限,新葉可替換枯黃老葉,例如:楓樹通常在秋末冬初落葉,隔年再長出新葉。

雙子葉植物的典型葉,外形可分為葉片、葉柄和托葉三部分,但有些植物的葉缺少葉柄或托葉;單子葉植物不具托葉,葉柄多呈鞘狀包在莖上,稱為葉鞘(圖2-13)。

葉柄或葉鞘連接莖和葉片,具支持與運輸功能,還可調整葉的角度與位置,以便獲得更多日照的機會。而托葉則是在葉柄基部兩側的突起,功能不明顯。多數植物的托葉只在葉芽剛伸長時可見,葉成熟時就脫落了。
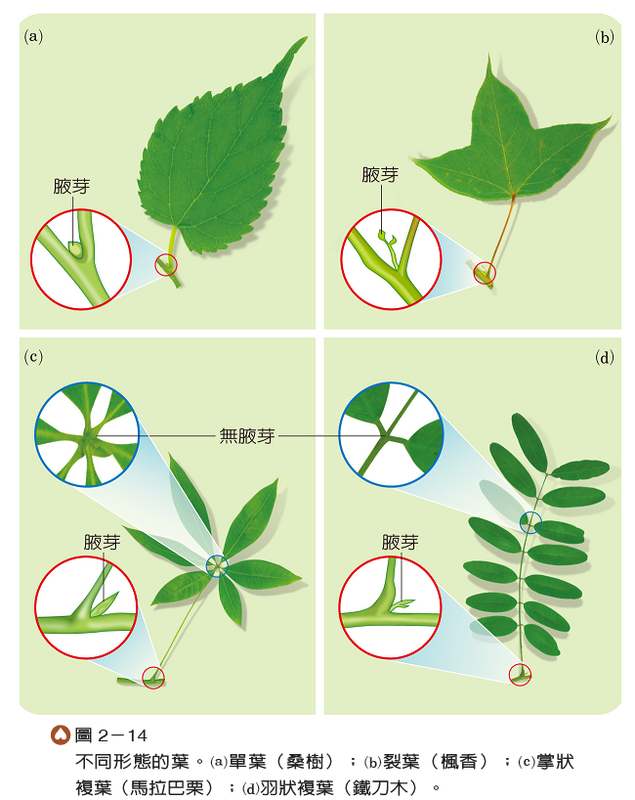
葉片多為扁平狀,但其外形輪廓則呈多樣化。葉片完整的葉,稱為單葉;有些單葉的葉片末端常裂成數片,但基部仍相連,又稱裂葉。有些葉片是由2枚以上分離的小葉所構成,稱為複葉,例如:掌狀複葉是由數枚小葉著生同一處,與葉柄相接;而羽狀複葉則是葉片由排列如羽毛狀的小葉所構成(圖2-14)。雙子葉植物的葉可分為單葉及複葉兩大類,單子葉植物則多為單葉。
葉片多為扁平狀,但其外形輪廓則呈多樣化。葉片完整的葉,稱為單葉;有些單葉的葉片末端常裂成數片,但基部仍相連,又稱裂葉。有些葉片是由2枚以上分離的小葉所構成,稱為複葉,例如:掌狀複葉是由數枚小葉著生同一處,與葉柄相接;而羽狀複葉則是葉片由排列如羽毛狀的小葉所構成(圖2-14)。雙子葉植物的葉可分為單葉及複葉兩大類,單子葉植物則多為單葉。
葉脈是葉上常見的明顯特徵,通常有一條最大的主脈貫穿葉片中央。葉脈的分布方式有網狀脈與平行脈,雙子葉植物通常為網狀脈,單子葉植物則多為平行脈(圖2-15)。

葉在莖上著生的情形稱為葉序,適當的排列可使植物在生長過程中,每一片葉都能接受到日照,不會被其他葉遮蔽。常見的葉序有互生、對生和輪生三種類型(圖2-16)。每個節上只著生1 枚葉者,稱為互生;著生2枚葉者,稱為對生;著生3枚葉以上者,稱為輪生。

點圖看常見葉序
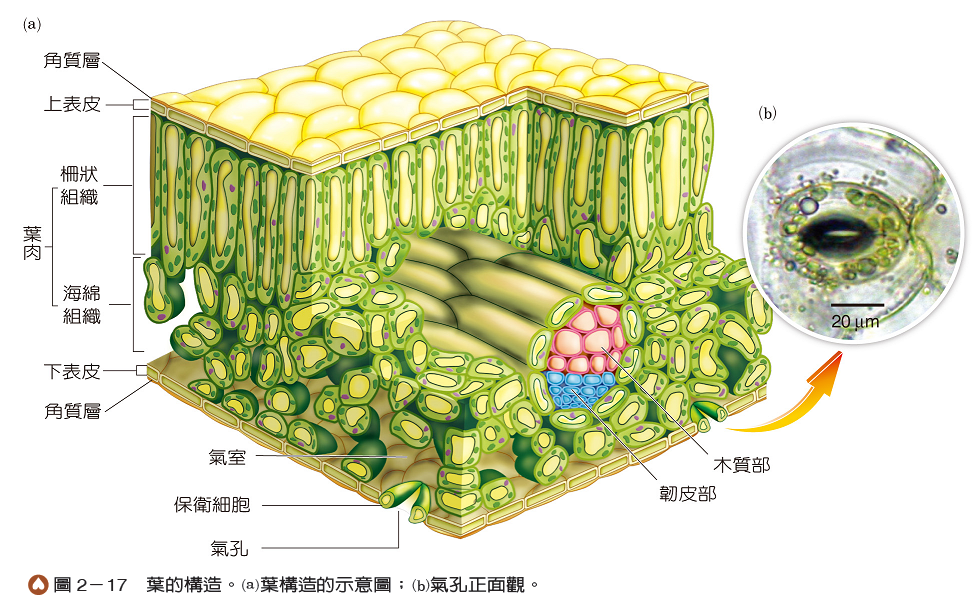
葉的構造由外而內包括表皮、葉肉和葉脈三個部分(圖2-17),此組成在雙子葉與單子葉植物間差異不大。葉的表皮通常只有一層細胞,可分為上表皮及下表皮,主要由表皮細胞和保衛細胞構成。表皮有角質層覆蓋,可減少水分的蒸散,並具有保護作用。保衛細胞通常呈腎形,兩兩成對,具有葉綠體,細胞壁的厚薄不均勻,內側壁較厚而外側壁較薄,兩細胞間的小孔稱為氣孔,是氣體進出的門戶(圖2-17)。此外,在不同植物中,葉的上、下表皮組成可能會有不同,例如:氣孔大多分布於下表皮,以減少直接照光而加速水分蒸散;又如表皮上可能有不同形態的毛或不同厚度的蠟質覆蓋,增加對葉片的保護。
葉肉位於上、下表皮之間,由具葉綠體的薄壁細胞所構成,是行光合作用的主要場所。有些植物的葉肉依細胞形狀與排列可分為兩型,其中一型的細胞多呈長橢圓形,靠近上表皮且與表皮細胞垂直,細胞排列較整齊,稱為柵狀組織;而另一型的細胞形狀不一,靠近下表皮且排列不整齊,細胞間有許多空隙,稱為海綿組織(圖2-17)。此外,在接近氣孔處,葉肉細胞常圍出一個大型空腔稱為氣室,是氣體暫存與交換的空間。
葉脈即是分布於葉肉中的維管束,包含木質部和韌皮部,木質部靠近上表皮側,韌皮部靠近下表皮側(圖2-17);具有支持平展的葉片及運輸水分和養分的功能。
葉肉位於上、下表皮之間,由具葉綠體的薄壁細胞所構成,是行光合作用的主要場所。有些植物的葉肉依細胞形狀與排列可分為兩型,其中一型的細胞多呈長橢圓形,靠近上表皮且與表皮細胞垂直,細胞排列較整齊,稱為柵狀組織;而另一型的細胞形狀不一,靠近下表皮且排列不整齊,細胞間有許多空隙,稱為海綿組織(圖2-17)。此外,在接近氣孔處,葉肉細胞常圍出一個大型空腔稱為氣室,是氣體暫存與交換的空間。
葉脈即是分布於葉肉中的維管束,包含木質部和韌皮部,木質部靠近上表皮側,韌皮部靠近下表皮側(圖2-17);具有支持平展的葉片及運輸水分和養分的功能。

有些植物的葉在形態或構造上不同於一般的葉,能適應特殊的生存環境。常見的變態葉包括:仙人掌的「刺」可減少水分散失,並兼具防禦的作用;九重葛的苞葉顏色鮮豔,可吸引昆蟲造訪,協助傳粉;豌豆葉的捲鬚可纏繞於接觸物上,協助植物體攀爬;捕蠅草的捕蟲葉可藉由葉的閉合來困住小型昆蟲,並分泌酵素分解屍體,再吸收含氮養分(圖2-18)。


植物從土壤中吸收水及無機鹽,藉由光合作用合成有機養分。植物體內的維管束在各部位間形成連通的管道,以運送水、無機鹽及有機養分至個體各部位,提供生長所需。

水和無機鹽由根吸收後,經維管束中的木質部由莖往上運輸至葉或其他生殖器官,以單向運輸的方式進行。
根為植物吸收水和無機鹽的主要部位,通常幼根的吸收能力較老根佳,尤其是成熟部的根毛區。根毛是表皮細胞的突起,可增加根與土壤顆粒的接觸面積,故可大幅提高吸收的效率。土壤中的無機鹽常會溶於水而形成離子狀態,如硝酸根(NO3-)等,須藉由表皮細胞膜上運輸蛋白的協助,以促進性擴散及主動運輸的方式被根吸收;水則可因根的內、外濃度差而滲透進入根中。
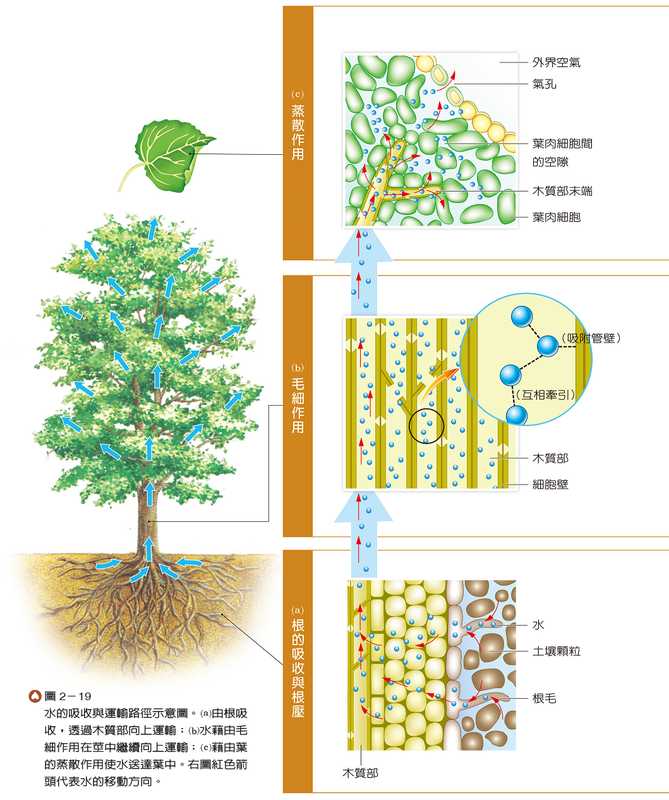
由根持續吸收的水和無機鹽,在經過皮層、內皮到達木質部後,逐漸在根中形成根壓,可推擠木質部中的水分向上運輸至莖(圖2-19(a))。由於水分子會吸附在木質部管壁上,加上水分子間互相牽引,故產生毛細作用,使水分沿著木質部繼續向上運輸(圖2-19(b))。無機鹽因溶於水中,亦會隨著水分運輸至各部位。
水分的運送除了有根壓的向上推擠以及木質部內毛細作用的協助外,主要還需水分自氣孔蒸散所產生的拉力,才能將莖部的水分子持續牽引至葉中,以供應葉肉細胞行光合作用(圖2-19(c)),因此,蒸散作用是植物體內水分向上運輸的主要動力。一般而言,植物體自根部所吸收的水分,90% 以上會經由氣孔蒸散到大氣中。
根為植物吸收水和無機鹽的主要部位,通常幼根的吸收能力較老根佳,尤其是成熟部的根毛區。根毛是表皮細胞的突起,可增加根與土壤顆粒的接觸面積,故可大幅提高吸收的效率。土壤中的無機鹽常會溶於水而形成離子狀態,如硝酸根(NO3-)等,須藉由表皮細胞膜上運輸蛋白的協助,以促進性擴散及主動運輸的方式被根吸收;水則可因根的內、外濃度差而滲透進入根中。
由根持續吸收的水和無機鹽,在經過皮層、內皮到達木質部後,逐漸在根中形成根壓,可推擠木質部中的水分向上運輸至莖(圖2-19(a))。由於水分子會吸附在木質部管壁上,加上水分子間互相牽引,故產生毛細作用,使水分沿著木質部繼續向上運輸(圖2-19(b))。無機鹽因溶於水中,亦會隨著水分運輸至各部位。
水分的運送除了有根壓的向上推擠以及木質部內毛細作用的協助外,主要還需水分自氣孔蒸散所產生的拉力,才能將莖部的水分子持續牽引至葉中,以供應葉肉細胞行光合作用(圖2-19(c)),因此,蒸散作用是植物體內水分向上運輸的主要動力。一般而言,植物體自根部所吸收的水分,90% 以上會經由氣孔蒸散到大氣中。

相較於水及無機鹽在木質部內的單向運輸,有機養分主要在維管束的韌皮部內可進行向上或向下的雙向運輸,其方向還會依各部位對養分的需求不同而改變。
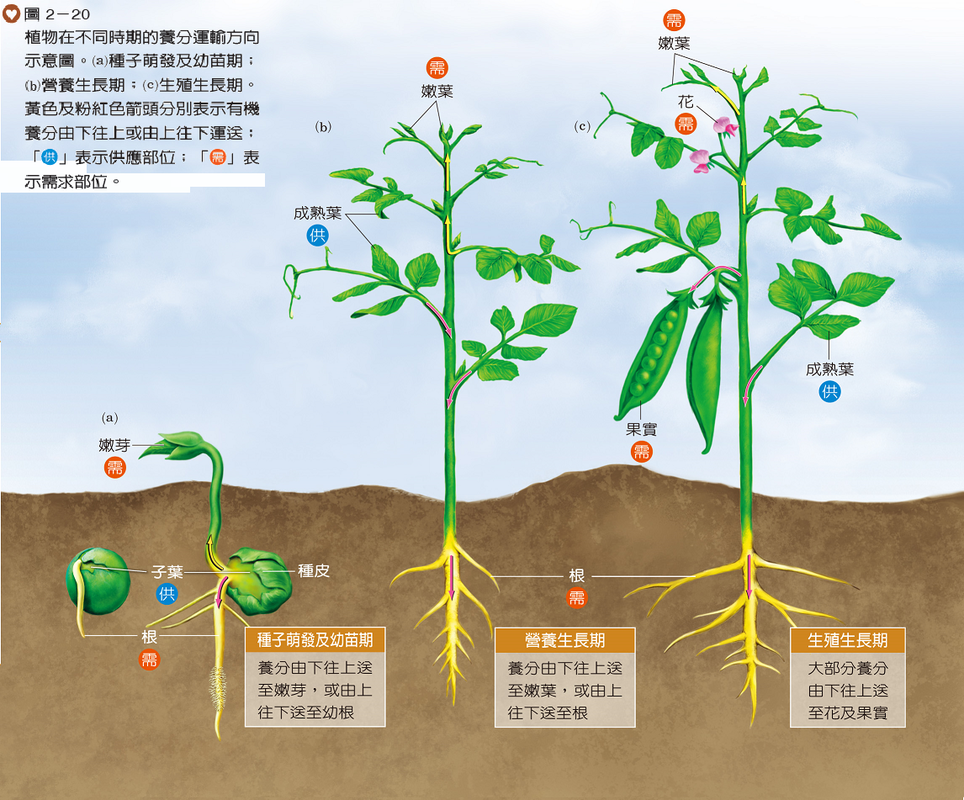
葉肉細胞行光合作用產生的有機養分,會先由葉脈收集,再經由莖往上運至頂端生長區供其利用,或往下運至根部利用並儲存。因此,植物體的成熟葉是提供有機養分的主要構造,相當於供應部位;而消耗養分或將剩餘養分儲存起來的構造,則相當於需求部位,例如:生長中的嫩芽、果實及根等。然而,種子、塊莖或儲藏根是供應部位抑或需求部位,則視其生長時期而定(圖2-20)。
葉肉細胞行光合作用產生的有機養分,會先由葉脈收集,再經由莖往上運至頂端生長區供其利用,或往下運至根部利用並儲存。因此,植物體的成熟葉是提供有機養分的主要構造,相當於供應部位;而消耗養分或將剩餘養分儲存起來的構造,則相當於需求部位,例如:生長中的嫩芽、果實及根等。然而,種子、塊莖或儲藏根是供應部位抑或需求部位,則視其生長時期而定(圖2-20)。